Полиплоидную породу тутового шелкопряда вывел

Получение полиплоидов тутового шелкопряда
Как были получены полиплоиды тутового шелкопряда?
Полиплоиды тутового шелкопряда были получены Б.Л. Астауровым с использованием метода температурного воздействия, который заключался в активации неоплодотворенных яиц путём погружения их в воду 46 °С на 18 мин с последующим охлаждением в течение 5–10 мин в воде комнатной температуры и содержанием их до наступления диапаузы при температуре 15–17 °С и относительной влажности 80–95 %. Это вызывало в неоплодотворенном яйце активацию развития, а тепловой шок подавлял редукционное деление и единственное остающееся эквационное деление созревания с самого начала обеспечивает диплоидность партеногенетического развития и генетическое тождество партеногенетических потомков – всегда самок – с матерью. Одним из оригинальных направлений исследований Б.Л. Астаурова была разработка «мужского» партеногенеза, или андрогенеза – способа получения только самцов. Для получения андрогенеза свежеоплодотворенные яйца подвергались также сильному прогреву, который выводил из строя чувствительный к прогреву ядерный аппарат ооцита и исключал из развития хромосомы матери. Ядро зародыша включало только отцовские хромосомы, внесенные спермиями. Развитие шло на основе материнской цитоплазмы и отцовского ядра. Андрогенетические особи были диплоидны, так как возникали в результате слияния ядер двух спермиев, и только самцы.
Партеногенез часто сопровождается появлением полиплоидных особей, а температурный партеногенез дал возможность получать их направленно. В 1940 г. Б.Л. Астауров заметил, что крупные ооциты, встречающиеся у партеногенетических самок, тетраплоидны. Путем термоактивации неоплодотворенных тетраплоидных яиц были получены тетраплоидные самки, способные как к партеногенетическому размножению, так и к размножению путем скрещивания. Так были получены аутотетраплоидные клоны. При скрещивании тетраплоидных самок с диплоидными самцами были получены триплоидные самки и самцы, которые оказались стерильными. В дальнейшем был найден путь разведения даже таких бесплодных триплоидных самок при температурном партеногенезе. Искусственный температурный партеногенез был осуществлён у Bombyx mori, у дикого шелкопряда Bombyx mandarina, у дубового шелкопряда Antheraea pernyi GuerMen., а также у межвидового гибрида Bombyx mori × Bombyx mandarina.
Источник
Полиплоидия
Полиплодия (от греческих слов polyploos — многократный и eidos — вид) — наследственное изменение, заключающееся в кратком увеличении числа наборов хромосом в клетках. Дети всегда похожи на обоих родителей. Происходит это потому, что в каждой их клетке находятся два набора хромосом, два комплекта генов — один материнский и один отцовский.
Такой двойной, или диплоидный (от греческих слов diploos — двойной и eidos — вид), набор хромосом типичен для живой природы. Он достаточен для преемственности поколений.
Но в некоторых тканях диплоидных организмов в процессе их развития появляются клетки, в которых 4, 8 или гораздо больше наборов хромосом. Такие клетки называют полиплоидными, а сам процесс — соматической полиплоидией (от греческого слова soma — тело).
Такая частичная полиплоидизация клеток некоторых тканей очень широко распространена, она свойственна всем изученным классам животных и растений. Например, у млекопитающих много полиплоидных клеток находят в печени, сердце, среди пигментных клеток и др.
Другое явление — генеративная полиплоидия, исходно свойственная организмам или искусственно создаваемая при оплодотворении. В этом случае все клетки организма полиплоидные.

Полиплоидия у гиацинта:
1 — гаплоидное пыльцевое зерно, содержащее в ядре 12 хромосом; 2 — диплоидное, содержащее 24 хромосомы; 3 — тетраплоидное — 48 хромосом.

Полиплоидия у ржи.
Справа — более крупные семена у растения-полиплоида.
Такой вариант полиплоидии наиболее свойствен растениям, особенно высшим.
Для полиплоидных растений обычно характерны крупные размеры. Избыток хромосом повышает их устойчивость к болезням и многим повреждающим воздействиям, например к радиации: при повреждении одной или даже двух сходных (гомологичных) хромосом остаются другие такие же совершенно целые.
Полиплоидные особи жизнеспособнее диплоидных. Многие виды растений полиплоидные. Вероятно, так же эволюционировали и некоторые животные. Примером могут служить некоторые черви, насекомые, рыбы и др.
Человек давно использует полиплоидию для выведения высокопродуктивных сортов сельскохозяйственных растений. Не так давно, до начала нашего века, это делалось бессознательно: просто размножали самые крупные экземпляры, дающие много зерна или же особенно крупные плоды. Отбором лучших растений закрепляли признак, нужный человеку.
С появлением генетики выяснилось, что такие гиганты — природные полиплоиды и, следовательно, их отбор — это выделение полиплоидного сорта из предкового, диплоидного вида.
Тогда полиплоиды стали создавать. Есть вещество колхицин, задерживающее деление клеток: число хромосом перед делением удваивается, как обычно, но клетка не делится, и в ней получается 4 набора хромосом. Воздействуя раствором колхицина на семена, можно получить полиплоидное растение. Задержать деление клетки можно также рентгеновским облучением, нагревом и некоторыми другими воздействиями. Можно воздействовать на гаметы и получить зиготу с умноженнным числом хромосом, которое сохранится во всех ее потомках — соматических клетках. У растений, которые размножаются и вегетативным путем (см. Размножение), можно получить полиплоидное потомство от природного или выведенного полиплоида.
Около 80% современных культурных растений — полиплоиды. Среди них хлебные злаки, овощные и плодовые культуры, многие ягодные, цитрусовые, некоторые технические и лекарственные растения. Среди сортов декоративных растений также немало полиплоидов.
Советские ученые вывели триплоидную свеклу, отличающуюся от обычной не только крупными размерами корнеплодов, но и повышенной их сахаристостью, а также устойчивостью к болезням. Выведена полиплоидная гречиха, гораздо более урожайная, чем исходные, диплоидные сорта. Возможно получение межвидовых полиплоидных гибридов, например ржи и пшеницы, капусты и редьки.
Экспериментально полученные полиплоиды животных — большая редкость. Так, советскому ученому-генетику Б. Л. Астаурову методом межвидовой гибридизации удалось получить полиплоидную форму тутового шелкопряда, продуцента шелка. Ученые вывели полиплоидных рыб, а в последнее время и птиц, например кур. Однако внедрение полиплоидных пород животных в практику сельского хозяйства — дело будущего.
Источник
Бабочка — тутовый шелкопряд

Из маленького бугорка под нижней губой гусеницы выделяется клейкое вещество, которое при соприкосновении с воздухом тут же застывает и превращается в шелковую нить. Нить очень тонкая, но выдерживает вес до 15 граммов.
Все современные домашние животные и культурные растения произошли от диких видов. Не обошлось в хозяйстве и без насекомого — бабочки тутового шелкопряда. За четыре с половиной тысячелетия селекционной работы удалось вывести породы, дающие шёлк разных цветов, а длина непрерывной нити с одного кокона может достигать километра! Бабочка так изменилась, что теперь трудно сказать, кто был её диким предком. В природе тутовый шелкопряд не встречается — без человеческой опеки он гибнет.
Вспомним, что и многие другие гусеницы плетут кокон из шелковистых нитей, но только у тутового шелкопряда они обладают нужными нам свойствами. Из шёлковых нитей производят ткани, очень прочные и красивые; их используют в медицине — для зашивания ран и очистки зубов; в косметологии — для изготовления декоративной косметики, например теней. Несмотря на появление искусственных материалов, натуральные шёлковые нити по-прежнему используются очень широко.
Кому же первому пришло в голову соткать шёлковую ткань? Как гласит легенда, четыре тысячи лет тому назад в чашку с горячим чаем, который пила в своём саду китайская императрица, упал кокон шелкопряда. Пытаясь вытащить его, женщина потянула за торчащую шелковинку. Кокон стал разматываться, но ниточка всё не кончалась. Тогда-то сметливая императрица и поняла, что из таких волокон можно делать пряжу. Китайский император одобрил идею жены и повелел своим подданным выращивать тутовник (белую шелковицу) и разводить на нём гусениц шелкопряда. И по сей день шёлк в Китае называют именем этой правительницы, а саму её благодарные потомки возвели в ранг божества.
Чтобы получить из коконов бабочки прекрасный шёлк, приходилось немало потрудиться. Для начала коконы нужно собрать, отбраковать и, главное, размотать, для чего их опускали в кипяток. Далее нить укрепляли серицином — шёлковым клеем, который затем удалялся кипящей водой или горячим мыльным раствором.

Вылупляющаяся бабочка размягчает кокон щелочной слюной и выбирается наружу.
Перед окрашиванием нить вываривали и отбеливали. Красили её растительными пигментами (плодами гардении, корнями морены, дубовыми желудями), либо минеральными (киноварью, охрой, малахитом, свинцовыми белилами). И только потом ткали пряжу — вручную или на ткацком станке.
Уже за полторы тысячи лет до нашей эры одежда из шёлковых тканей была в Китае обычна. В других азиатских странах и у древних римлян шёлк появился только в III веке до нашей эры — и стоил он тогда баснословно дорого. Но технология изготовления удивительной ткани ещё долгие века оставалась тайной для всего мира, ведь попытка вывезти шелкопряда за пределы Китайской империи каралась смертной казнью. Природа шёлка представлялась европейцам загадочной и волшебной. Одни верили, что шёлк производят гигантские жуки, другие считали, что в Китае земля мягкая, как шерсть, и поэтому после полива её можно использовать для получения шёлковых тканей.
Открылся же секрет шёлка в IV веке нашей эры, когда китайская принцесса преподнесла дар своему жениху — королю Малой Бухары. То были яйца шелковичного червя, которые невеста тайно вывезла с родины, спрятав в своей причёске. Примерно тогда же секрет шёлка стал известен японскому императору, но здесь шелководство некоторое время было монополией одного лишь императорского дворца. Потом производство шёлка освоили и в Индии. А оттуда с двумя монахами, поместившими яйца шелкопряда в полые ручки своих посохов, те попали в Византию. В XII—XIV веках шелководство расцвело в Малой Азии, Испании, Италии и во Франции, а в XVI появилось в южных провинциях России.

Куколка тутового шелкопряда
Однако и после того как европейцы научились разводить шелкопряда, большую часть шёлка продолжали доставлять из Китая. По Великому шёлковому пути — сети дорог, идущих с востока на запад, — его везли во все страны мира. Шёлковые наряды оставались предметом роскоши, шёлк служил и обменной валютой.
Как же живёт маленькая белая бабочка — «шёлковая королева»? Размах её крыльев — 40—60 миллиметров, но в результате многолетнего культивирования бабочки утратили способность к полёту. Ротовой аппарат не развит, потому что взрослая особь не питается. Завидным аппетитом отличаются только личинки. Их выкармливают листьями тутового дерева (шелковицы). При кормлении же другими растениями, которые гусеницы «соглашаются» есть, качество волокна ухудшается. На территории нашей страны представители семейства настоящих шелкопрядов, к которому относится и тутовый шелкопряд, в природе встречаются только на Дальнем Востоке.

Разные породы тутового шелкопряда дают шёлк разного цвета.
Гусеницы тутового шелкопряда выводятся из яиц, кладка которых покрыта плотной оболочкой и называется греной. В шелководческих хозяйствах грену помещают в специальные инкубаторы, где поддерживаются необходимые температура и влажность. Через несколько дней на свет появляются маленькие, трёхмиллиметровые личинки тёмно-бурого цвета, покрытые пучками длинных волос.
Вылупившихся гусениц переносят на специальную кормовую этажерку со свежими листьями шелковицы. После нескольких линек малютки подрастают до восьми сантиметров, а их тела становятся белыми и почти голыми.
Готовая к окукливанию гусеница перестаёт питаться, и тогда по соседству с ней помещают древесные прутья, на которые она тотчас же переходит. Удерживаясь брюшными ножками за один из прутьев, гусеница закидывает голову то вправо, то назад, то влево и прикладывается своей нижней губой с «шёлковым» бугорком к различным местам прута.

Гусениц выкармливают листьями тутового дерева.
Вскоре вокруг неё образуется довольно густая сеть из шёлковой нити. Но это ещё только основа будущего кокона. Затем «мастерица» переползает в центр каркаса и начинает завивать нить: выпуская её, гусеница быстро вертит головой. Неутомимая ткачиха трудится над коконом около четырёх суток! А после замирает в своей шёлковой колыбели и превращается там в куколку. Примерно через 20 дней из куколки выходит бабочка. Она размягчает кокон своей щелочной слюной и, помогая себе ножками, с трудом выбирается наружу, чтобы приступить к поиску партнёра для продолжения рода. После спаривания самка откладывает 300—600 яиц.
Однако не каждой гусенице дано превратиться в бабочку. Большую часть коконов отправляют на фабрику для получения шёлка-сырца. Один центнер таких коконов даёт примерно девять килограммов шёлковой нити.
Интересно, что гусеницы, из которых в дальнейшем получаются самцы, — работники более старательные, их коконы плотнее, а значит, нить в них длиннее. Учёные научились регулировать пол бабочек, повышая выход шёлка при его промышленном производстве.
Такова история маленькой белой бабочки, которая прославила Древний Китай и заставила весь мир поклоняться своему великолепному продукту.
Ольга Тимохова, кандидат биологических наук
Источник
Полиплоидия у животных
Полиплоидия широко представлена у растений.
Это связано с тем, что у растений весьма распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Очевидно, существуют и другие причины, способствующие полиплоидии у растений. К таким причинам, по-видимому, могут относиться специфические для гибридных растений ядерно-плазменные отношения.
Особенно большие препятствия для сохранения полиплоидных клеток и их воспроизведения в поколениях возникают у раздельно — полых организмов, имеющих гетерогаметный и гомогаметный пол. Установлено, что у раздельнополых животных полиплоидия встречается редко. У тех животных, у которых половое размножение заменено партеногенезом, полиплоидия может осуществляться почти так же, как у растений.
О полиплоидных рядах у животных вообще говорить еще преждевременно. В этом отношении могут быть названы пока единичные примеры, известные у аскариды, земляных червей, амфибий и некоторых др. Так, исследованиями С. Мюльдаля установлено, что в семействе земляных червей (Lumbricidae) имеются полиплоидные ряды с разными основными числами: 11, 16, 17, 18 и 19 хромосом, в некоторых из них найдены даже декаплоиды. Все эти полиплоиды размножаются обычно партеногенетически. Полиплоидные земляные черви оказываются более крупными, чем ближайшие родственные им диплоидные виды. Более известны у животных двучленные полиплоидные ряды, т. е. диплоиды (2n) и тетраплоиды (4n). Такие ряды обнаружены у некоторых насекомых и амфибий. В настоящее время получены автополиплоидные особи у тутового шелкопряда, тритона, аксолотля и даже у млекопитающих (мышь, кролик).
Явление партеногенетического развития неоплодотворенных яиц довольно часто встречается у птиц. У индюшек найдены некоторые линии, у которых яйца развиваются партеногенетически даже еще до инкубации. В таких линиях около 80% зародышевых дисков яиц девственны. Большинство из них является диплоидными, но встречаются также и гаплоиды.
У автотетраплоидов шелковичного червя (Bombyx mori) самки плодовиты, а самцы стерильны. Причиной этого является то, что у самцов (гомогаметный пол) в профазе мейоза образуются поливаленты, в силу чего возникают гаметы с анеуплоидным числом хромосом. У самок (гетерогаметный пол) поливаленты не образуются, а если и образуются, то не препятствуют распределению хромосом вследствие отсутствия кроссинговера; мейоз у них протекает более нормально.
У аксолотля было получено несколько поколений автотетраплоидных и триплоидных самок. Самки амфибий, являясь генетически гетерогаметным полом, в случае автотетраплоидии оказываются частично или полностью плодовитыми. Самцы, являясь гомогаметным полом, оказываются полностью стерильными. То же самое имеет место у тутового шелкопряда. Интересно также отметить, что триплоидные самки аксолотля оказываются более жизнеспособными и плодовитыми, чем тетраплоидные.
У млекопитающих (в частности, у домашней мыши и кролика), по-видимому, возможно получение полиплоидных яйцеклеток. Этого удается достигнуть путем воздействия тепловым или холодовым шоком на оплодотворенное яйцо, в котором такие воздействия задерживают второе деление созревания. В результате яйцеклетка оказывается с диплоидным набором хромосом. После слияния материнского диплоидного ядра с мужским гаплоидным пронуклеусом образуется триплоидная зигота (мейотическая полиплоидия). Этот механизм образования триплоидных зигот является общим для насекомых, амфибий и млекопитающих. Кроме этого способа, существует другой, а именно — подавление тепловым шоком (45,5°) первого деления дробления, что дает возможность получать тетраплоидные зиготы.
Таким образом, причинами образования триплоидии у животных могут быть:
- полиандрия, когда два сперматозоида сливаются с гаплоидным ядром яйцеклетки;
- полигиния, когда один сперматозоид сливается с двумя гаплоидными ядрами в яйцеклетке;
- анеугамия, когда один сперматозоид сливается с диплоидной (не созревшей) яйцеклеткой.
Недавно (1963 г.) у кур был обнаружен случай спонтанной триплоидии по аутосомам с хромосомной формулой 3А + XX.
Эта курица имела нормальную жизнеспособность, но ее правая гонада была рудиментарной, а левая — мозаичной — частью мужеской и частью женской.
У млекопитающих также возникают триплоидные зиготы благодаря полиандрии и полигинии. Например, полиандрия у крыс встречается с частотой 1,2—3,2% от числа оплодотворенных яйцеклеток. Такая же частота обнаружена у золотистых хомячков, полевок и мышей. Однако триплоидные эмбрионы у мышей доживают только до половины беременности. По мнению некоторых авторов, случаи триплоидии могут иметь место и у человека. Триплоидия встречается в цикле размножения насекомых с партеногенетическим развитием. Особенно успешное изучение распространения триплоидии в природных популяциях возможно у тех насекомых, у которых имеются гигантские хромосомы в слюнных железах. На этих хромосомах удается точно установить гомологичность хромосом.
Все указанные примеры касаются автополиплоидии. Получение аллополиплоидов у животных до недавнего времени считалось неразрешимой проблемой. Однако совсем недавно Б. Л. Астаурову с сотрудниками удалось искусственно создать первый аллополиплоид от межвидового гибрида шелкопрядов Bombyx mori X В. mandarina. У обоих этих видов n = 28 хромосомам.
При синтезировании аллотетраплоида использовался метод искусственного партеногенеза. Вначале были получены партеногенетические автополиплоиды В. mori — 4 n, 6 n. Все эти особи оказались женского пола и были плодовиты. Затем произвели скрещивание партеногенетических самок В. mori (4n) с самцами другого вида В. mandarina (2n). В потомстве от такого скрещивания появлялись аллотриплоидные самки 2n В. mori + 1 n В. mandarina. Эти самки, стерильные в обычных условиях, размножались путем партеногенеза. При этом партеногенетически иногда возникали аллогексаплоидные самки 4n В. mori + 2n В. mandarina. В потомстве от скрещивания этих самок с диплоидными самцами В. mandarina (2n) были отобраны формы обоего пола с удвоенным набором хромосом каждого вида 2n В. mori +2n В. mandarina — аллотетраплоиды, или амфидиплоиды.
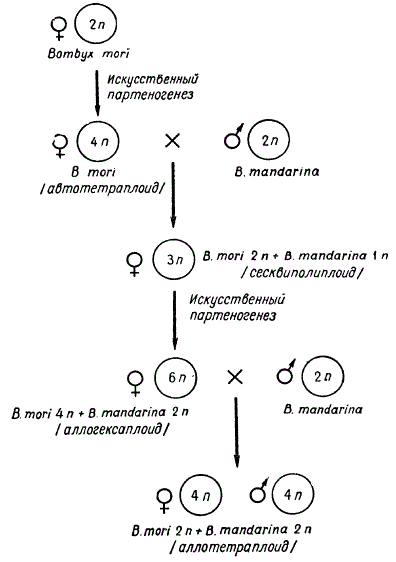
Схема получения аллотетраплоида у Bombyx
Если гибрид 1n В. mori + 1n В. mandarina был бесплодным, то аллотетраплоид оказался плодовитым и при разведении внутри себя дал плодовитое потомство. Плодовитость этих форм была невысокой, как и следует ожидать, в силу отсутствия селекции, но к настоящему времени воспроизведено уже шесть поколений аллотетраплоидов. С помощью полиплоидии, таким образом, удалось синтезировать новую форму шелкопряда. В. Н. Верейская, исследуя мейоз у самцов аллотетраплоидов, пришла к выводу, что значительная стерильность у них зависит от образования поливалентов, приводящих к возникновению анеуплоидных нежизнеспособных сперматозоидов. А образование поливалентов у аллотетраплоида объясняется высокой гомологией хромосом указанных двух видов.
Выше мы отметили, что полиплоидия у раздельнополых животных является довольно редким событием. Для объяснения причины такого явления Г. Мёллером было высказано два предположения:
1) препятствием сохранения полиплоидии у раздельнополых животных является изменение нормального соотношения половых хромосом и аутосом в зиготе, что ведет к нарушению полового генного баланса и возникновению стерильности особей;
2) так как вероятность встречи редуцированных гамет двух полов тетраплоидов очень мала, то тетраплоидная бисексуальная форма обычно скрещивается с диплоидной, и поэтому гибриды почти всегда оказываются триплоидными и, следовательно, стерильными.
Недавно Б. Л. Астауров обратил внимание еще на одну причину препятствия полиплоидии у бисексуальных форм — практически полное бесплодие одного из полов при полиплоидии, вследствие образования поливалентных комплексов. Таким полом является, как правило, гомогаметный, у которого совершается кроссинговер, ведущий к анеуплоидии гамет.
Однако не следует думать, что полиплоидия в мире животных в принципе ограничена. Она довольно широко распространена в соматических клетках ряда тканей многоклеточных организмов, т. е. в тех случаях, где нет синапсиса хромосом и нормально текущего кроссинговера, а также редукции числа хромосом.
В последние годы усиленно изучается явление полиплоидии у простейших. Оказывается, что в ряде случаев полиплоидия у них играла существенную роль в изменении циклических превращений ядра и возникновении ядерного дуализма, т. е. наличия макро- и микронуклеусов. Установлено, что генетически активным ядром, определяющим фенотип инфузории, является макронуклеус, а микронуклеус прямо не влияет на фенотип; он является лишь Передатчиком наследственной информации при половом процессе. В исследованиях Ю. И. Полянского и его сотрудников (И. Б. Майкова и др.), а также ряда зарубежных авторов установлено, его макронуклеус инфузорий является автополиплоидным; в нем насчитываются сотни геномов. При таком умножении наборов идентичных геномов митоз утрачивает свое значение как механизм равного распределения наследственного материала между дочерними клетками при бесполом размножении инфузорий. Предполагается, что на этой основе митоз в эволюции мог замениться амитотическим делением макронуклеусов. Наличие большого числа идентичных геномов даже при амитозе всегда обеспечивает попадание нескольких геномов в каждую дочернюю клетку. Это очень правдоподобное объяснение раскрывает важную проблему происхождения амитотического деления. Такое объяснение позволяет пересмотреть представление об амитозе как о филогенетически более древней форме деления ядра по сравнению с митозом. С данной точки зрения амитоз мог возникнуть как вторичное явление на основе высокой плоидности ядра.
В настоящее время большинство генетиков придерживается той точки зрения, что в эволюции животных основную роль играла все же не полиплоидия, а межхромосомные и внутрихромосомные перестройки. И эта точка зрения сейчас наиболее оправдана. У млекопитающих, как правило, большое число хромосом. Возможно, что наблюдающаяся у них изменчивость соматического числа хромосом обязана хромосомным мутациям типа транслокации. Эволюция кариотипов посредством фрагментации хромосом кажется для них наиболее вероятной. Однако пути эволюции организмов разнообразны, и механизмы, ею выработанные, приурочены к конкретной эволюции каждого класса, отряда, рода и вида.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Источник







