Вывел полиплоидный гибрид тутового шелкопряда

Получение полиплоидов тутового шелкопряда
Как были получены полиплоиды тутового шелкопряда?
Полиплоиды тутового шелкопряда были получены Б.Л. Астауровым с использованием метода температурного воздействия, который заключался в активации неоплодотворенных яиц путём погружения их в воду 46 °С на 18 мин с последующим охлаждением в течение 5–10 мин в воде комнатной температуры и содержанием их до наступления диапаузы при температуре 15–17 °С и относительной влажности 80–95 %. Это вызывало в неоплодотворенном яйце активацию развития, а тепловой шок подавлял редукционное деление и единственное остающееся эквационное деление созревания с самого начала обеспечивает диплоидность партеногенетического развития и генетическое тождество партеногенетических потомков – всегда самок – с матерью. Одним из оригинальных направлений исследований Б.Л. Астаурова была разработка «мужского» партеногенеза, или андрогенеза – способа получения только самцов. Для получения андрогенеза свежеоплодотворенные яйца подвергались также сильному прогреву, который выводил из строя чувствительный к прогреву ядерный аппарат ооцита и исключал из развития хромосомы матери. Ядро зародыша включало только отцовские хромосомы, внесенные спермиями. Развитие шло на основе материнской цитоплазмы и отцовского ядра. Андрогенетические особи были диплоидны, так как возникали в результате слияния ядер двух спермиев, и только самцы.
Партеногенез часто сопровождается появлением полиплоидных особей, а температурный партеногенез дал возможность получать их направленно. В 1940 г. Б.Л. Астауров заметил, что крупные ооциты, встречающиеся у партеногенетических самок, тетраплоидны. Путем термоактивации неоплодотворенных тетраплоидных яиц были получены тетраплоидные самки, способные как к партеногенетическому размножению, так и к размножению путем скрещивания. Так были получены аутотетраплоидные клоны. При скрещивании тетраплоидных самок с диплоидными самцами были получены триплоидные самки и самцы, которые оказались стерильными. В дальнейшем был найден путь разведения даже таких бесплодных триплоидных самок при температурном партеногенезе. Искусственный температурный партеногенез был осуществлён у Bombyx mori, у дикого шелкопряда Bombyx mandarina, у дубового шелкопряда Antheraea pernyi GuerMen., а также у межвидового гибрида Bombyx mori × Bombyx mandarina.
Источник
Гибридизация. Эксперимент Менделя. Полиплоидия
Гибридизация – процесс скрещивания гибридов.
Гибридизация подразделяется на внутривидовую, то есть скрещивание одного вида. А также по систематическим группам, то есть скрещивание геномов разных видов.
Для первого поколения гибридов всегда характерен гетерозис (увеличение жизнеспособности гибридов вследствие полученных различных наборов генов для гибридизации). Если отдаленная гибридизация, то гибрид стерилен (бесплоден).
Гибридизация невозможна, если разный хромосомный набор у видов. Если у гибрида происходит геномная мутация, вызывающая процесс удвоения хромосом, процесс мейоза возможен. При этом приобретается способность к размножению, но полностью теряется возможность скрещивания с родительской формой вида. Так же межвидовые гибриды могут размножаться вегетативным путём.
В природе существуют растения, гибриды которых могли получиться таким путём.
Существуют виды пшеницы с 14, 28, 42 хромосомами. Виды роз с 14, 28, 42 и 56 хромосомами. У фиалок существуют хромосомные наборы кратные 6, которые варьируются от 12 до 54.
Не менее трети видов цветковых растений мы имеем благодаря гибридизации.
В пример можно привести эксперимент Грегора Менделя(1822-1884).
Мендель в своём эксперименте проводил искусственную гибридизацию, а выведенные виды скрещивал между собой. В этом эксперименте учёный изучил 7 признаков у 20 тысяч гибридов второго поколения. Учёный выбрал удачный для исследования растение – горох. Горох в норме – самоопылитель и на нём легко проводить искусственную гибридизацию. В своём опыте он не только точно описал процессы наследуемых признаков, также использовал математический анализ.
Мендель показал, что у всех растений первого поколения от родителей наследуется только один доминантный признак. При скрещивании пурпурного и белого цветка гороха в ходе эксперимента взошли растения только с пурпурным цветом. При скрещивании жёлтого гороха с зелёным у всех потомков семена были жёлтыми. В случае с гладкими и морщинистыми семенами гороха получились гладкие семя. Проявление одного признака от одного из родителей учёный назвал его доминантным. Эта закономерность сформировала первый закон Менделя.
Во втором поколении при скрещивании гибридов первого поколения у каждого четвёртого гибрида проявлялся рецессивный признак. Среди трёх растений гороха с пурпурными цветками одно было с белыми. На три стручка с жёлтыми семенами приходился один зёленый. На три растения с гладкими семенами приходился один с морщинистыми. Явление скрещивания гетерозиготных особей приводит к образованию потомства, где одна часть несёт доминантный признак, а другая рецессивный, называют «расщеплением». Это сформировало второй закон Менделя.
При скрещивании растений жёлтых и гладких семян гороха(доминантные признаки растения) с растениями, которые были зелёные и морщинистые семена(рецессивные признаки), во втором поколении произвели расщепление в пропорции 3:1(3 части жёлтых семян и 1 часть зелёных семян) и 3:1(3 части гладких семян и 1 часть морщинистых семян). Расщепление по одному признаку шло независимо от другого. Это явление сформировало третий закон Менделя.

Так же у Менделя есть ещё один известный эксперимент с кошкой.
У родителей с разными наследственными признаками потомки первого поколения получат признаки только одного родителя. К примеру – при скрещивании чёрного кота с рыжей кошкой котята будут иметь чёрный окрас(доминантный признак). Это дало образование закона единообразия.
При скрещивании разных особей, родители которых имели разные доминантные признаки, возможно, получат признаки предшествующего поколения. Например, при скрещивании белой кошки и рыжего кота мы получим котят окраса обоих родителей и одного рыжего котёнка. Это дало начало закона расщепления признаков.
При скрещивании особей, у которых разнятся несколько признаков, мы получим следующее: при скрещивании рыжей гладкошерстной кошки и белого длинношерстного кота получим разнообразное потомство, среди которых будут копии родителей, рыжие длинношерстные и белые гладкошерстные. Это сформировало закон независимого наследования.
Так же в селекции широко применяют метод полиплоидии. Полиплоидия – увеличение числа хромосом. Полиплоидию применяют при создании сортов пшеницы, ржи, овса, картофеля, хлопчатника, плодовых, декоративных и других культур. Полиплоидные растения так же могут возникать и в результате естественных мутаций. Применяют методы искусственного получения полиплоидов, воздействуют мутагенами, в основном колхицином. Таким образом получают из диплоидных(2n) тетраплоидные(4n) хромосомные наборы. Полиплоидные растения отличаются от обычных высокой урожайностью, размерами, активным синтезом органических веществ. Использование полиплоидии позволило селекционерам вывести особо ценные сорта сахарной свеклы, гречихи, ржи, фасоли и многих других культур.

В селекции животных метод полиплоидии применяется крайне редко. Самым ярким исключением является эксперимент Бориса Астаурова(1904-1974). Он вывел полиплоидные гибриды тутового шелкопряда, размножающегося партеногенезом.
Отдалённая гибридизация позволяет в одном организме совместить признаки разных видов или родов. Получение таких видов из-за нескрещиваемости родителей и стерильности(бесплодия) гибридов делает его сложным. Для восстановления плодовитости таких гибридов генетик Георгий Дмитриевич Карпеченко предложил использовать метод полиплоидии. В гибриде капусты и редьки он, убедившись в её стерильности, выделил полиплоидную форму, которая оказалась плодоносной.
Центры происхождения культурных растений
Н. И. Вавилов установил восемь центров происхождения культурных растений:
1) восточноазиатский – родина сои, проса, гречихи, многие плодовитые и овощные культуры;
2) южноазиатский тропический – родина риса, сахарного тростника, цитрусовых, многие овощные культуры;
3) юго-западная – пшеница, рожь, бобовые, лён, конопля, морковь, виноград;
4) переднеазиатский – мягкая пшеница, ячмень, овёс;
5) среднеземноморский – родина капусты, свёклы, маслин;
6) абиссинский – родина твёрдой пшеницы, сорго, бананов, кофе;
7) центральноамериканский – кукуруза, какао, тыква, табак, хлопчатник;
8) южноамериканский – картофель, ананас, хинное дерево.
Источник
Полиплоидия у животных
Полиплоидия широко представлена у растений.
Это связано с тем, что у растений весьма распространены гермафродитизм (самоопыление), апомиксис (партеногенез) и вегетативное размножение. Очевидно, существуют и другие причины, способствующие полиплоидии у растений. К таким причинам, по-видимому, могут относиться специфические для гибридных растений ядерно-плазменные отношения.
Особенно большие препятствия для сохранения полиплоидных клеток и их воспроизведения в поколениях возникают у раздельно — полых организмов, имеющих гетерогаметный и гомогаметный пол. Установлено, что у раздельнополых животных полиплоидия встречается редко. У тех животных, у которых половое размножение заменено партеногенезом, полиплоидия может осуществляться почти так же, как у растений.
О полиплоидных рядах у животных вообще говорить еще преждевременно. В этом отношении могут быть названы пока единичные примеры, известные у аскариды, земляных червей, амфибий и некоторых др. Так, исследованиями С. Мюльдаля установлено, что в семействе земляных червей (Lumbricidae) имеются полиплоидные ряды с разными основными числами: 11, 16, 17, 18 и 19 хромосом, в некоторых из них найдены даже декаплоиды. Все эти полиплоиды размножаются обычно партеногенетически. Полиплоидные земляные черви оказываются более крупными, чем ближайшие родственные им диплоидные виды. Более известны у животных двучленные полиплоидные ряды, т. е. диплоиды (2n) и тетраплоиды (4n). Такие ряды обнаружены у некоторых насекомых и амфибий. В настоящее время получены автополиплоидные особи у тутового шелкопряда, тритона, аксолотля и даже у млекопитающих (мышь, кролик).
Явление партеногенетического развития неоплодотворенных яиц довольно часто встречается у птиц. У индюшек найдены некоторые линии, у которых яйца развиваются партеногенетически даже еще до инкубации. В таких линиях около 80% зародышевых дисков яиц девственны. Большинство из них является диплоидными, но встречаются также и гаплоиды.
У автотетраплоидов шелковичного червя (Bombyx mori) самки плодовиты, а самцы стерильны. Причиной этого является то, что у самцов (гомогаметный пол) в профазе мейоза образуются поливаленты, в силу чего возникают гаметы с анеуплоидным числом хромосом. У самок (гетерогаметный пол) поливаленты не образуются, а если и образуются, то не препятствуют распределению хромосом вследствие отсутствия кроссинговера; мейоз у них протекает более нормально.
У аксолотля было получено несколько поколений автотетраплоидных и триплоидных самок. Самки амфибий, являясь генетически гетерогаметным полом, в случае автотетраплоидии оказываются частично или полностью плодовитыми. Самцы, являясь гомогаметным полом, оказываются полностью стерильными. То же самое имеет место у тутового шелкопряда. Интересно также отметить, что триплоидные самки аксолотля оказываются более жизнеспособными и плодовитыми, чем тетраплоидные.
У млекопитающих (в частности, у домашней мыши и кролика), по-видимому, возможно получение полиплоидных яйцеклеток. Этого удается достигнуть путем воздействия тепловым или холодовым шоком на оплодотворенное яйцо, в котором такие воздействия задерживают второе деление созревания. В результате яйцеклетка оказывается с диплоидным набором хромосом. После слияния материнского диплоидного ядра с мужским гаплоидным пронуклеусом образуется триплоидная зигота (мейотическая полиплоидия). Этот механизм образования триплоидных зигот является общим для насекомых, амфибий и млекопитающих. Кроме этого способа, существует другой, а именно — подавление тепловым шоком (45,5°) первого деления дробления, что дает возможность получать тетраплоидные зиготы.
Таким образом, причинами образования триплоидии у животных могут быть:
- полиандрия, когда два сперматозоида сливаются с гаплоидным ядром яйцеклетки;
- полигиния, когда один сперматозоид сливается с двумя гаплоидными ядрами в яйцеклетке;
- анеугамия, когда один сперматозоид сливается с диплоидной (не созревшей) яйцеклеткой.
Недавно (1963 г.) у кур был обнаружен случай спонтанной триплоидии по аутосомам с хромосомной формулой 3А + XX.
Эта курица имела нормальную жизнеспособность, но ее правая гонада была рудиментарной, а левая — мозаичной — частью мужеской и частью женской.
У млекопитающих также возникают триплоидные зиготы благодаря полиандрии и полигинии. Например, полиандрия у крыс встречается с частотой 1,2—3,2% от числа оплодотворенных яйцеклеток. Такая же частота обнаружена у золотистых хомячков, полевок и мышей. Однако триплоидные эмбрионы у мышей доживают только до половины беременности. По мнению некоторых авторов, случаи триплоидии могут иметь место и у человека. Триплоидия встречается в цикле размножения насекомых с партеногенетическим развитием. Особенно успешное изучение распространения триплоидии в природных популяциях возможно у тех насекомых, у которых имеются гигантские хромосомы в слюнных железах. На этих хромосомах удается точно установить гомологичность хромосом.
Все указанные примеры касаются автополиплоидии. Получение аллополиплоидов у животных до недавнего времени считалось неразрешимой проблемой. Однако совсем недавно Б. Л. Астаурову с сотрудниками удалось искусственно создать первый аллополиплоид от межвидового гибрида шелкопрядов Bombyx mori X В. mandarina. У обоих этих видов n = 28 хромосомам.
При синтезировании аллотетраплоида использовался метод искусственного партеногенеза. Вначале были получены партеногенетические автополиплоиды В. mori — 4 n, 6 n. Все эти особи оказались женского пола и были плодовиты. Затем произвели скрещивание партеногенетических самок В. mori (4n) с самцами другого вида В. mandarina (2n). В потомстве от такого скрещивания появлялись аллотриплоидные самки 2n В. mori + 1 n В. mandarina. Эти самки, стерильные в обычных условиях, размножались путем партеногенеза. При этом партеногенетически иногда возникали аллогексаплоидные самки 4n В. mori + 2n В. mandarina. В потомстве от скрещивания этих самок с диплоидными самцами В. mandarina (2n) были отобраны формы обоего пола с удвоенным набором хромосом каждого вида 2n В. mori +2n В. mandarina — аллотетраплоиды, или амфидиплоиды.
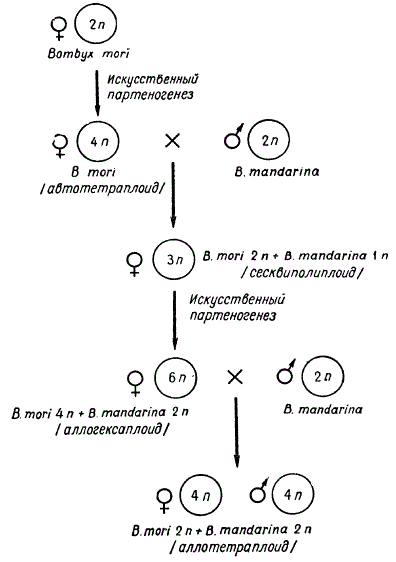
Схема получения аллотетраплоида у Bombyx
Если гибрид 1n В. mori + 1n В. mandarina был бесплодным, то аллотетраплоид оказался плодовитым и при разведении внутри себя дал плодовитое потомство. Плодовитость этих форм была невысокой, как и следует ожидать, в силу отсутствия селекции, но к настоящему времени воспроизведено уже шесть поколений аллотетраплоидов. С помощью полиплоидии, таким образом, удалось синтезировать новую форму шелкопряда. В. Н. Верейская, исследуя мейоз у самцов аллотетраплоидов, пришла к выводу, что значительная стерильность у них зависит от образования поливалентов, приводящих к возникновению анеуплоидных нежизнеспособных сперматозоидов. А образование поливалентов у аллотетраплоида объясняется высокой гомологией хромосом указанных двух видов.
Выше мы отметили, что полиплоидия у раздельнополых животных является довольно редким событием. Для объяснения причины такого явления Г. Мёллером было высказано два предположения:
1) препятствием сохранения полиплоидии у раздельнополых животных является изменение нормального соотношения половых хромосом и аутосом в зиготе, что ведет к нарушению полового генного баланса и возникновению стерильности особей;
2) так как вероятность встречи редуцированных гамет двух полов тетраплоидов очень мала, то тетраплоидная бисексуальная форма обычно скрещивается с диплоидной, и поэтому гибриды почти всегда оказываются триплоидными и, следовательно, стерильными.
Недавно Б. Л. Астауров обратил внимание еще на одну причину препятствия полиплоидии у бисексуальных форм — практически полное бесплодие одного из полов при полиплоидии, вследствие образования поливалентных комплексов. Таким полом является, как правило, гомогаметный, у которого совершается кроссинговер, ведущий к анеуплоидии гамет.
Однако не следует думать, что полиплоидия в мире животных в принципе ограничена. Она довольно широко распространена в соматических клетках ряда тканей многоклеточных организмов, т. е. в тех случаях, где нет синапсиса хромосом и нормально текущего кроссинговера, а также редукции числа хромосом.
В последние годы усиленно изучается явление полиплоидии у простейших. Оказывается, что в ряде случаев полиплоидия у них играла существенную роль в изменении циклических превращений ядра и возникновении ядерного дуализма, т. е. наличия макро- и микронуклеусов. Установлено, что генетически активным ядром, определяющим фенотип инфузории, является макронуклеус, а микронуклеус прямо не влияет на фенотип; он является лишь Передатчиком наследственной информации при половом процессе. В исследованиях Ю. И. Полянского и его сотрудников (И. Б. Майкова и др.), а также ряда зарубежных авторов установлено, его макронуклеус инфузорий является автополиплоидным; в нем насчитываются сотни геномов. При таком умножении наборов идентичных геномов митоз утрачивает свое значение как механизм равного распределения наследственного материала между дочерними клетками при бесполом размножении инфузорий. Предполагается, что на этой основе митоз в эволюции мог замениться амитотическим делением макронуклеусов. Наличие большого числа идентичных геномов даже при амитозе всегда обеспечивает попадание нескольких геномов в каждую дочернюю клетку. Это очень правдоподобное объяснение раскрывает важную проблему происхождения амитотического деления. Такое объяснение позволяет пересмотреть представление об амитозе как о филогенетически более древней форме деления ядра по сравнению с митозом. С данной точки зрения амитоз мог возникнуть как вторичное явление на основе высокой плоидности ядра.
В настоящее время большинство генетиков придерживается той точки зрения, что в эволюции животных основную роль играла все же не полиплоидия, а межхромосомные и внутрихромосомные перестройки. И эта точка зрения сейчас наиболее оправдана. У млекопитающих, как правило, большое число хромосом. Возможно, что наблюдающаяся у них изменчивость соматического числа хромосом обязана хромосомным мутациям типа транслокации. Эволюция кариотипов посредством фрагментации хромосом кажется для них наиболее вероятной. Однако пути эволюции организмов разнообразны, и механизмы, ею выработанные, приурочены к конкретной эволюции каждого класса, отряда, рода и вида.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Источник







